Estudios originales
← vista completaPublicado el 22 de junio de 2026 | http://doi.org/10.5867/medwave.2026.05.3191
Análisis mecanicista de los casos de COVID-19 en Chile durante el segundo semestre de 2020: un modelo SIR con tasa de transmisión dinámica
Mechanistic analysis of COVID-19 cases in Chile during the second half of 2020: an SIR model with dynamic transmission rate
Abstract
Introduction This article analyzes the prolonged trough phase in the epidemic curve associated with COVID-19 dynamics in Chile during the period July-December 2020, characterized by a relatively stable daily record of 1 000-2500 cases.
Methods Unlike traditional (, )-SIR models, with constant parameters and associated respectively with the transmission and removal rates in the infectious process, which predict unimodal behavior, we propose an approach based on Contagion Mechanics that incorporates a dynamic law for the transmission rate .
Results Using official data from the Department of Health Statistics and Information, we demonstrate how this approach quantitatively captures the observed stabilization, resulting from sustained adherence to non-pharmaceutical measures by the Chilean population. The model reveals that maintaining the infection rate below its intrinsic value required sustained collective effort, enabling controlled management of hospital demand during the pre-vaccination stage.
Conclusions Our results validate the usefulness of Contagion Mechanics in explaining complex epidemiological dynamics and offer new perspectives on the population response to prolonged health interventions.
Main messages
- Human behaviour plays a key role in the spread of disease, and in particular in the number of COVID-19 cases.
- This article presents a structurally simple mathematical model that provides a description reasonably consistent with data on the course of the COVID-19 pandemic in Chile.
- This work helps to rationalise decision-making by monitoring the dynamics without losing explanatory power.
- However, the model is not suitable for projections beyond the time period considered (July to December 2020).
Introduction
In general, the COVID-19 pandemic exhibited complex dynamics that challenged traditional mathematical modeling approaches across populations and observation levels [1,2]. In Chile, according to data from the Ministry of Health [3], following a first wave with a peak of 6938 daily cases reported in mid-June 2020, the epidemiological curve entered a phase of unusually prolonged stabilisation. As shown in Figure 1, from mid-July to the end of December 2020, daily confirmed cases remained predominantly between 1000 and 2500, forming an epidemiological ‘trough’. From the perspective of mathematical modeling, this behaviour contrasts with the typical unimodal patterns predictable using traditional SIR or SEIR (Susceptible-Exposed-Infected-Removed) models with a constant transmission rate [5,6]. This intermediate period, prior to the second wave recorded in early 2021, constitutes a unique case study for analysing the cause-and-effect relationship between the non-pharmaceutical mitigation measures implemented in the pre-vaccine era and the maintenance of that trough.
Daily number of COVID-19 cases reported in Chile by the WHO.
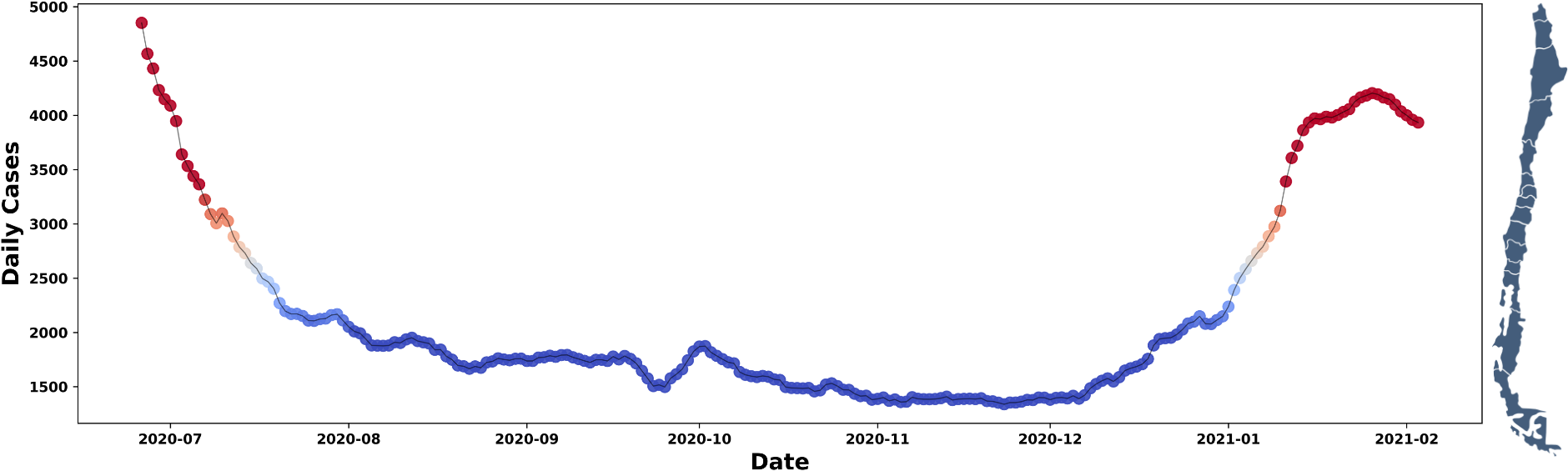
The colours indicate the magnitude: red (≥ 3000); orange (2501 to 2999); and in blue (≤ 2500) we highlight a prolonged trough with a minimum of 1338 cases on 23 November 2020.
WHO: World Health Organisation.
Source: compiled by the authors based on WHO records [4].
Indeed, traditional mathematical models, based on the classic SIR compartmental model and assuming a constant transmission rate, are inherently incapable of reproducing sequences of peaks and troughs. This is because they tend to predict unimodal incidence curves with sharp peaks [7,8,9]. As Chowell et al. point out, “simple epidemic growth models assume constant transmission rates and homogeneous mixing, which generally results in unimodal incidence curves that do not capture the complex resurgence patterns observed in historical pandemics” [10]. It is widely accepted that temporal variability in the transmission rate, driven by behavioural changes, public health interventions and population adherence to these, is fundamental to explaining such dynamics [11]. In turn, Flaxman et al. [12] demonstrated that non-pharmaceutical interventions, particularly lockdowns, significantly reduced transmission by substantially altering the effective reproduction number.
Broadly speaking, there are two strategic modeling approaches to incorporate this variability:
-
Imposing an explicit, a priori functional form for the transmission rate (for example, a function that decreases over time) [13,14,15,16].
-
Incorporating a dynamic law governing its evolution as a function of the system’s state variables [11,17].
Recently, the second approach has seen significant advances with the formulation of the mechanical theory of contagion [18,19,20]. An underlying objective of this article is to disseminate the language of the mechanical theory of contagion using a simple example represented by a basic model. Note that this theory provides a formal framework that allows the dynamics of the transmission rate to be derived from first principles, thereby avoiding ad hoc assumptions. These principles revolve around mechanisms of action and reaction associated with the loss and acquisition of intrinsic protective behaviours, linked to the implementation of pharmaceutical and non-pharmaceutical interventions [11,21,22]. Consequently, adherence to and compliance with these measures by the majority of the population influence the increase or decrease in the spread of the pathogen through social behaviour [23,24,25,26].
It is well documented that human behaviour is a key factor in fluctuations in the transmission rate [27,28,29]. The hypothesis put forward is that keeping the transmission rate below its natural or intrinsic level requires sustained behavioural effort on the part of the public. This led to a stabilisation which, in particular, allowed hospital demand to be kept generally below capacity, even taking into account capacity increases [30,31,32,33]. It should be noted that during this period, vaccines were not yet available [34].
In this context, our aim is to understand, from a general perspective, the behaviour of the infection rate in Chile during the period from July to December 2020, using the recent conceptual framework of the mechanical theory of contagion. We seek to quantify the collective effort required, in terms of reducing and stabilising the infection rate, to keep hospital demand at manageable levels. The results of the analysis not only validate the model’s explanatory power but also offer a mechanistic perspective on the population’s response to prolonged control measures in a critical scenario.
The article is structured into the following sections:
-
Methods, where the theoretical framework of the β-SIR model is developed, formalising the incorporation of a variable transmission rate governed by mechanistic-behavioural principles.
-
Results, in which a systematic numerical exploration is carried out to validate the model against data observed in Chile.
-
Discussions and conclusions, where the findings are summarised and future research directions are outlined.
Methods
We will consider a population affected by the spread of an infectious disease that follows the basic SIR compartmental epidemiological model. That is, each individual can be classified as susceptible (S), infectious (I) or recovered (R), and a unidirectional flow S → I → R is described by the system
Where
Assuming that, in human populations, a threat to life triggers a defensive response in individuals (in a similar way to what occurs in ecological communities, where prey seek shelter in the presence or under direct threat of a predator [35]), it is possible to consider the existence of a population fraction
Regarding
Let’s observe in (2) that
Thus, the change in the refugee population, estimated based on the balance between daily arrivals and departures, is given by
A key factor is understanding how this group of refugees affects the transmission rate
If we consider the fraction
Therefore, by deriving the identity in (4), we obtain
This equation corresponds to the reaction–replenishment dynamic law governing the transmission rate that Córdova-Lepe and Vogt-Geisse [20] refer to
Therefore, highlighting the dynamic nature of
With initial status
Introducing the concept of the ‘intrinsic reproduction number’, denoted by
Within the framework of contagion dynamics, introduced by Córdova-Lepe [18], the Second Law of Contagion concerns the balance between the strength of contagion
where, denoted by
Please note that we have highlighted the units
Since our focus is on the transient behaviour of model (6) (a very specific initial period associated with 2020), an analysis of its long-term behaviour is not of interest to us. However, it should be noted that if
Therefore, we observe that
This finding has significant epidemiological implications. The effective transmission rate never exceeds the intrinsic rate
Results
Depending on different estimated values of the intrinsic reproduction number
The initial conditions were derived from data reported by the Chilean Ministry of Health in its 10 July 2020 report [3]. In particular, the following were taken into account:
Scenario 1
Taking into account an average incubation period of
Trend in new and estimated COVID-19 cases, scenario 1.
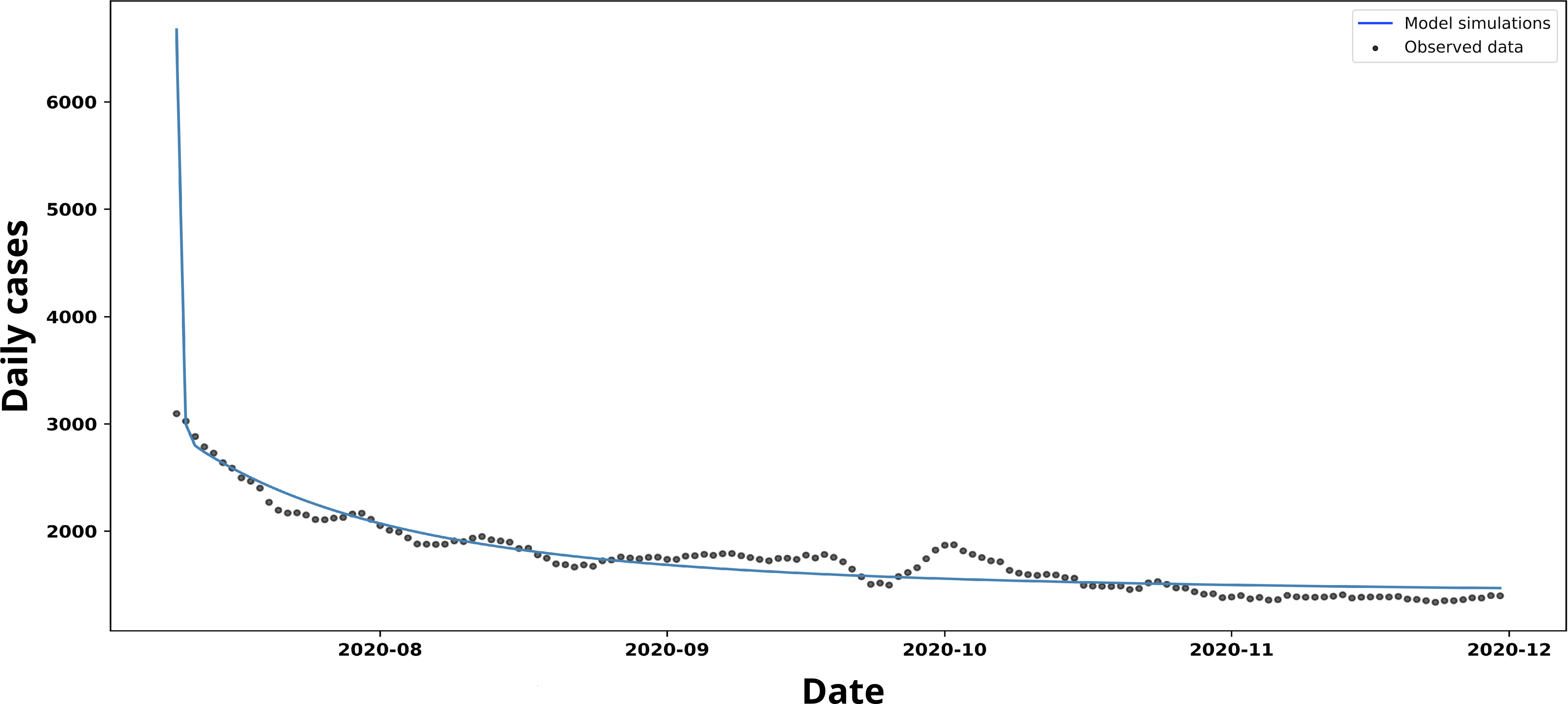
Source: Prepared by the authors based on the study’s results.
The fit based on the non-linear least squares method yielded the following values
Parameter tuning, scenario 1.
In (b), corresponding behaviour of the dynamic transmission loop tβ(t).
Source: Prepared by the authors based on the study results.
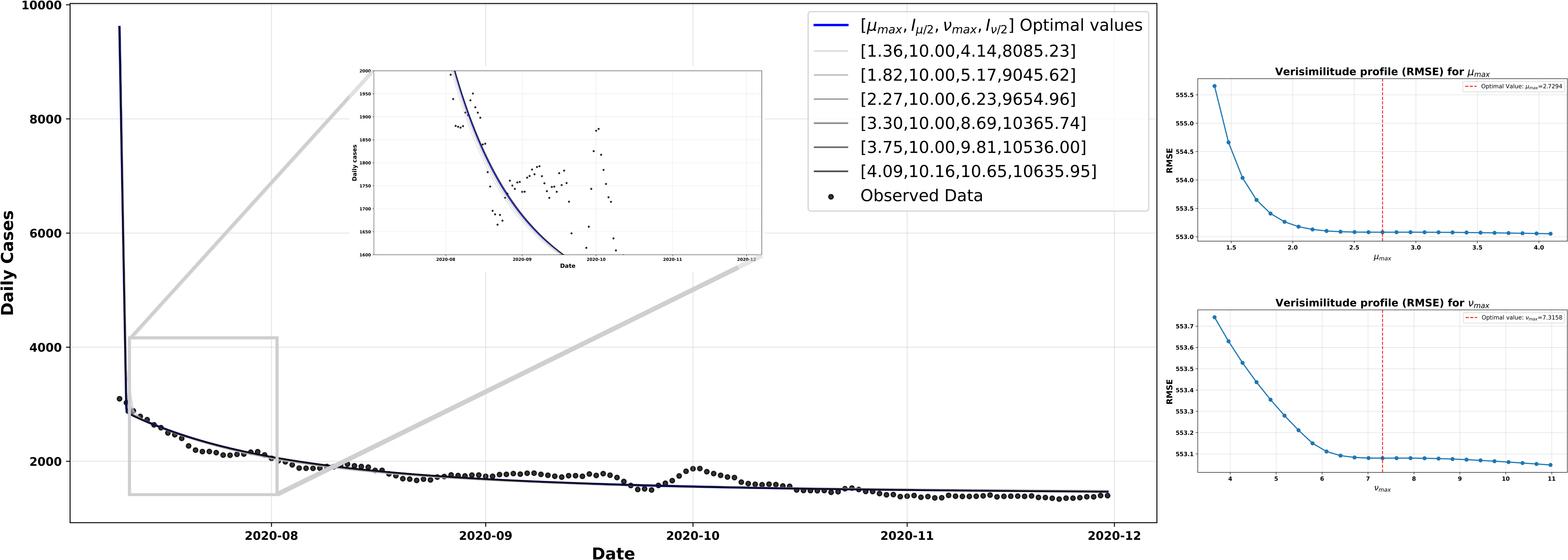
In Figure 3,
However, these adjusted values for protection and unprotection are difficult to identify in practice. As shown in the root mean square error (RMSE) curves in Figure 4, although both parameters can be estimated within the explored range, neither exhibits a clearly defined minimum. Rather than converging towards a clearly marked optimal value, the curves gradually flatten out and appear to settle at a local minimum.
Adjustment of protection and de-protection parameters, scenario 1.
RMSE: root mean square error.
Source: Prepared by the authors based on the study results.
This behaviour is consistent with what occurs during model fitting in this scenario: the parameters do not act independently. The epidemiological dynamics are non-linear and coupled, such that a change in any of the parameters alters the course of
The multiple configurations used for the simulations in Figure 4 were explored in a controlled manner by applying ±50% variations to the parameters
Scenario 2
For the simulation of scenario two, shown in Figure 5, an initial transmission rate was implemented
Trend in new and estimated COVID-19 cases, scenario 2.
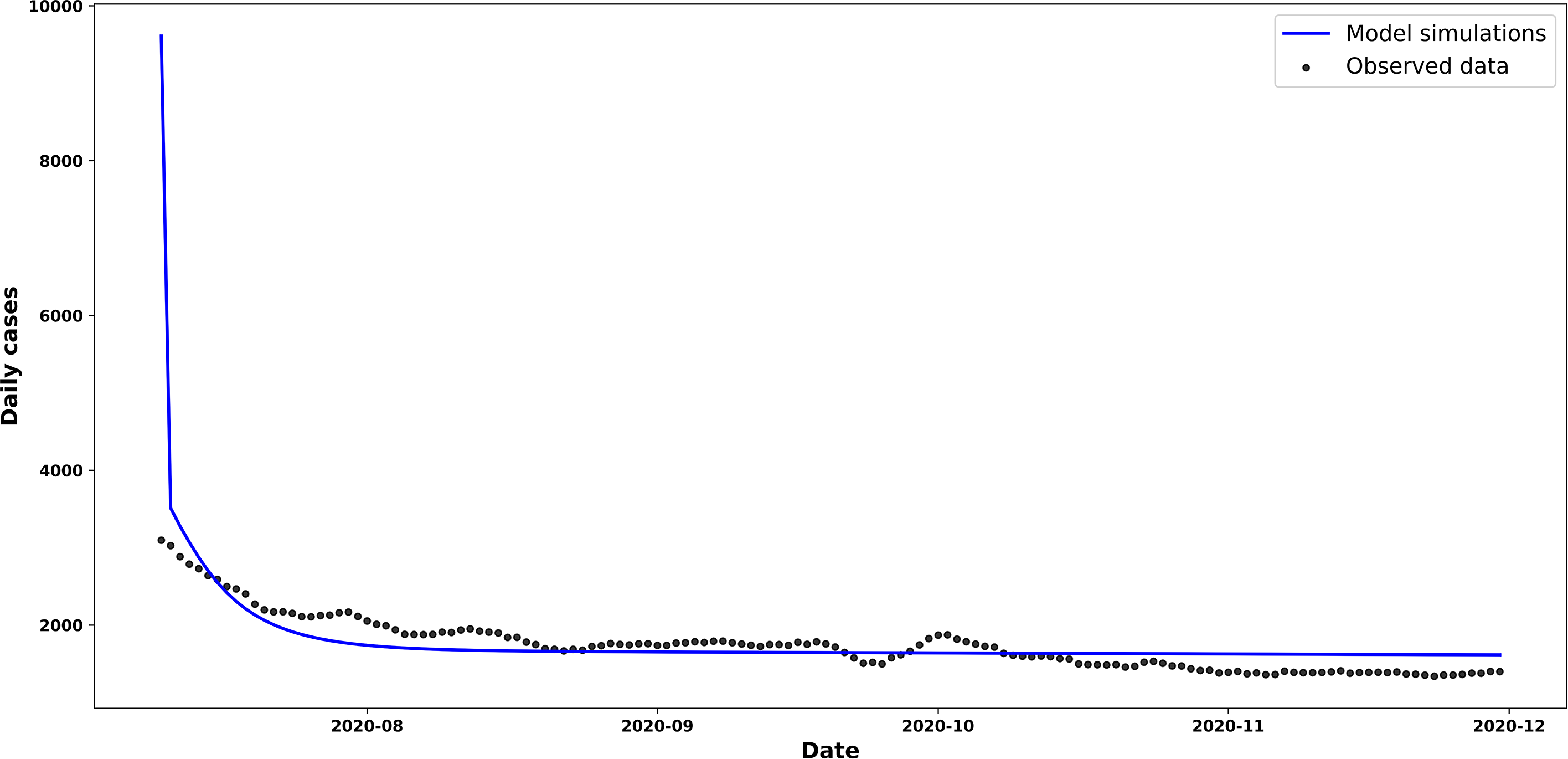
Source: prepared by the authors based on the study’s results.
The curve of new cases shown in Figure 5 follows a trend very similar to that simulated using the parameters of Scenario 1. However, the model slightly overestimates the number of cases towards the end of the period, whilst fitting the data quite accurately from the first few weeks onwards.
The non-linear least squares fit shown in Figure 6 yielded the values for the protection and unprotection parameters
Adjustment of protection and de-protection parameters, scenario 2.
RMSE: root mean square error.
Source: Prepared by the authors based on the study results.
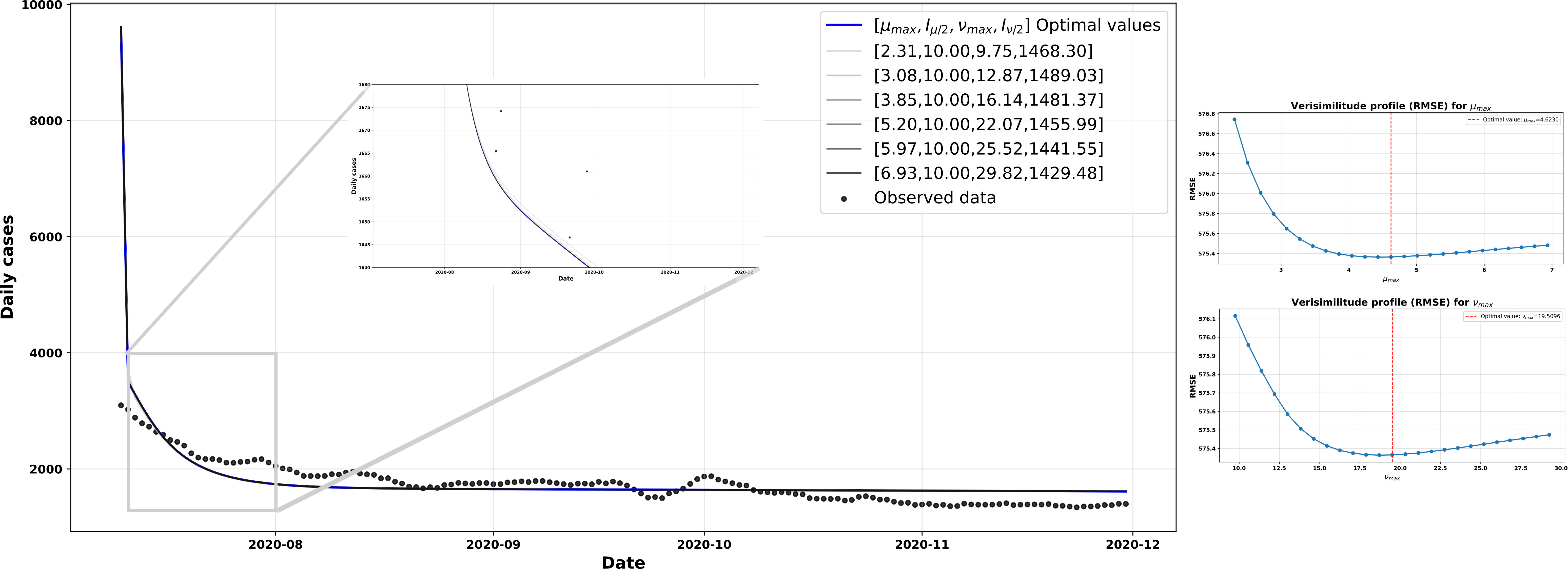
Scenario 3
In this scenario, we present the simulations based on the rates used and estimated by Canals et al [20]. In this study, the size of the Chilean population was taken as
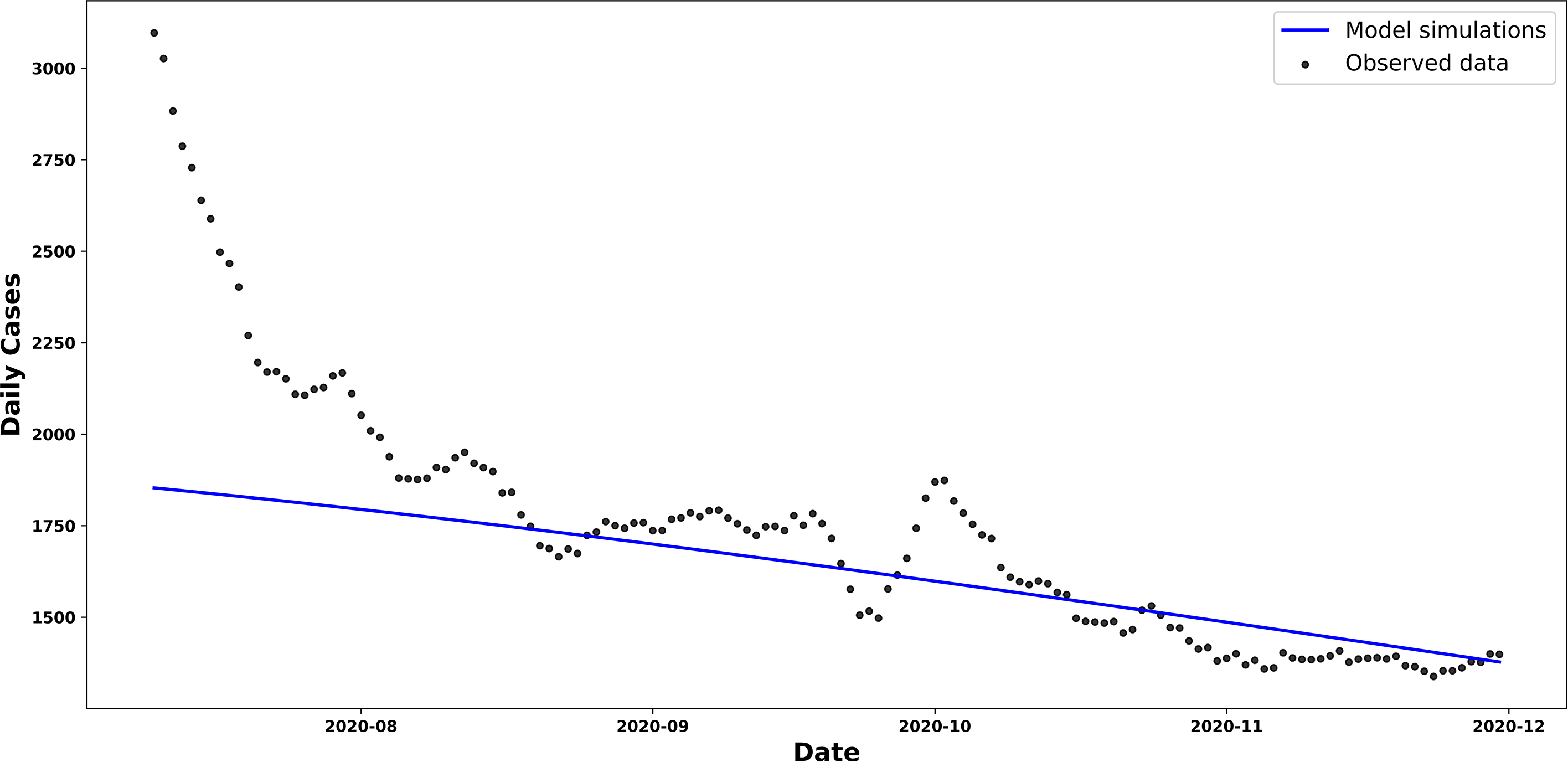
The curve of new cases shown in Figure 7 exhibits a linear trend that differs significantly from the simulated curves in the previous scenarios. It can be seen that the
Trend in new and estimated COVID-19 cases, scenario 3.
Comparative graph showing the trend in new COVID-19 cases in Chile and the trend in new cases estimated by our β-SIR model, as given by equation (6).
The protection parameters set for this scenario were
As for the adjusted vulnerability parameters, they were
Consequently, the protective effect provided by
The various configurations used for the simulations in Figure 8 were explored by applying ±50% variations to the parameters
The left-hand side shows model simulations obtained by shifting the optimal values of the parameters µmax and Vmax’ by 50%, compared with the observed data. On the right are the likelihood profiles based on the RMSE for µmax (top) and Vmax’ (bottom), where the red vertical line indicates the estimated optimal value for each parameter.
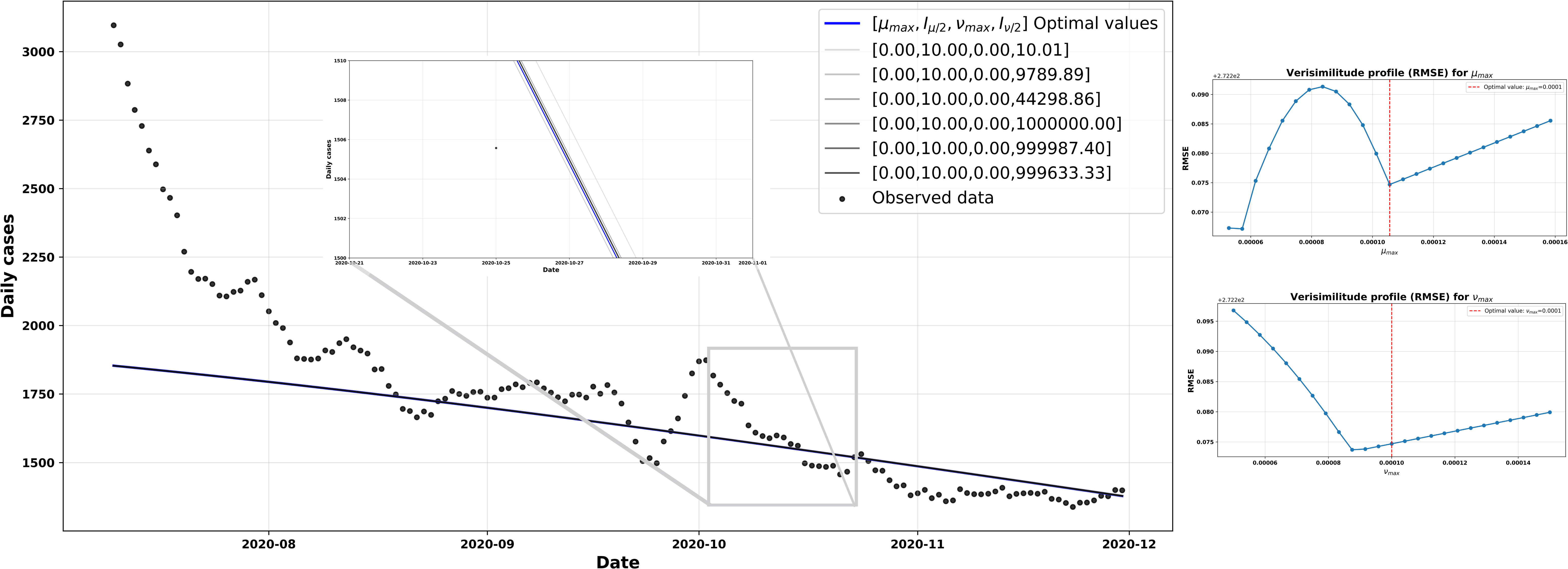
Source: Prepared by the authors of this study.
Scenario 4
In this final scenario, we expanded our analysis by explicitly incorporating the effect of the non-pharmaceutical measures implemented in Chile on the model’s dynamics. To do this, we used the composite stringency index. This indicator combines nine types of government interventions, including school and workplace closures, movement restrictions, and travel bans. This index ranges from 0 to 100, with higher values representing stricter policies, as reported by the World Health Organisation (WHO) [4].
The central idea was to allow the maximum protection parameter,
where the average stringency index was calculated over the period from 26 June 2020 to 30 January 2021, corresponding to the extended interval used in the simulation. Under this formulation, when the country adopts stricter measures, the protective effect incorporated into the model increases. Conversely, when measures are relaxed, this effect decreases. With this formulation, the initial conditions and parameters were selected
Rigor index.
Variation in the rigor index with respect to µmax.
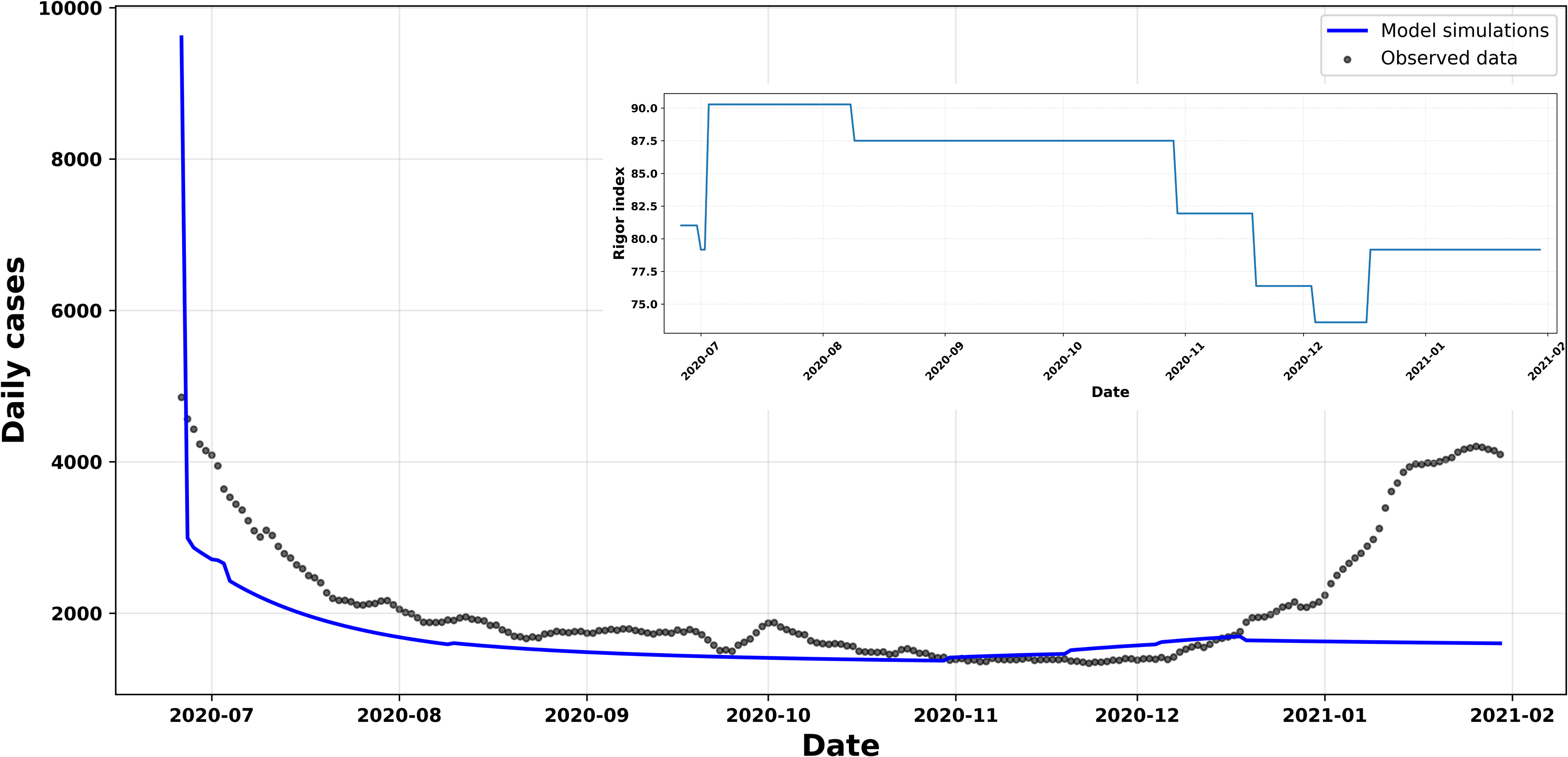
By incorporating the composite severity index into Figure 9, we observe that the parameters associated with the protection mechanism, in particular
Taken together, these results show that the model is capable of adequately capturing the influence of public policies on the population’s protective behaviour, whilst maintaining structural stability in its parameters throughout the adjustment.
Finally, we can observe in Table 1 that the model
Furthermore, the protective parameters applied to the transmission rate
The data suggest that these measures did not significantly impact the eradication of the disease. Chambon et al. [37] argue that the expected trajectory of the outbreak is influenced by the ‘unengaged’ or ‘non-compliant’ population in relation to non-pharmaceutical measures. Furthermore, they note that adherence to mitigation measures declines over time; and that factors such as pandemic fatigue, a perceived decrease in risk, and the normalisation of social contact limit the effectiveness of interventions.
The threshold value
Note that a common feature across all scenarios is the stabilisation of daily cases, a situation that persists as long as the external protective force
Discussion and conclusions
When faced with an epidemic curve, determining what constitutes a wave is not a settled matter, as there is no universal objective definition. This lack of consensus has direct consequences for detailed data analysis [38]. In this study, for the COVID-19 data in Chile during 2020, we have selected a period falling between the first two so-called waves. In a population of nearly twenty million people, the daily case curve shows relative stability which, at first glance (see Figure 1), forms a flat or trough-like section. We refer to what is commonly termed a trough, roughly bounded between early July (following a peak of 6,938 cases on 14 June [39]) and late December (prior to a new peak on 9 January 2021, with 4,956 cases). This period, associated with the dominant ‘Delta’ variant [40], shows a well-defined trough with a low of 1,352 cases on 22 November. This is of great interest for understanding the causes of this dynamic pattern of controlled or reduced transmission.
The literature suggests that these troughs may be due to a combination of factors, notably the depletion of susceptible individuals, seasonal changes, the effectiveness of control measures, and population behaviour. Ruling out the first two causes, we understand that, in the Chilean case, the observed stability is mainly associated with the implementation of health restrictions and a sufficiently alert population, willing to maintain, albeit to a decreasing extent, preventive behaviours. In this regard, the model with
It is possible to speculate that this trough began to disappear around January 2021, when figures rose again, probably due to behavioural factors associated with the end-of-year festivities, the start of the summer season and the announcement (on 16 December) of the arrival of 20,000 vaccine doses. This announcement materialised on the 24th of the same month, with the first person being vaccinated in the country [42].
The knowledge generated for decision-makers, most of whom belong to the medical community and the political sphere, is indispensable in the management of a high-risk infectious pandemic. For their part, mathematical models, given the phenomenon’s complexity, constitute essential tools. Hence the need to strengthen educational efforts in this direction, in order to better understand the course of transmission processes [43].
The aim of this article has been to present a structurally simple mathematical model—for example, one with a small number of compartments—yet capable of providing a description that is reasonably consistent with the data. Undoubtedly, even simple models play a significant role in the monitoring of epidemics, as they help to rationalise decision-making by tracking the dynamics without losing explanatory power [44]. In particular, this approach allows for an interpretative reading grounded in the language of the mechanics of infection, which introduces a Newtonian analogy based on concepts of force. From this perspective, non-pharmaceutical mitigation measures can be understood as a reaction force exerted by the population, the counterpart to the intrinsic cultural and environmental pressure driving a return to a natural state. However, the model is not viable for projections beyond the time interval considered. Indeed, after December 2020, it is no longer possible to assume that the system governing the dynamics remains a basic SIR model, as this assumption is unsustainable. This is because one must take into account the emergence of variants (non-homogeneous virulence and latency) and new structures arising from the multiplication of population compartments (for example, new infectious types), or the reduction in the susceptible population due to vaccination.
It should be noted that the usefulness or limitations of the mechanics of transmission—a theory which, in this paper, has been presented primarily as a descriptive and interpretative tool within a simplified empirical and theoretical context—depend on the modeler’s ability to transpose, by analogy, the physics of classical mechanics of multi-component systems into the language of transmission forces offered by the theory. An example of this application is in more complex epidemiological scenarios, such as the forces acting on transmission rates differentiated according to some compartmentalisation criterion (such as infectious types). However, for a more direct and comprehensive understanding of the scope and limits of the mechanics of contagion, the reader is invited to delve into the seminal publications [11,19,20] and the dynamics of the novel theory [18].
